by Vahe Andonians on May 22, 2023 9:14:46 AM
Artificial Neural Networks, in the broadest sense, mimic how our brain works. However, consequential simplifications result in contrasting AI and brain somewhat different behavior of Artificial Neural Networks compared to our brains.
In a series of articles, I highlight the differences between our brain and Artificial Neural Networks, focusing only on the parts that matter most. I am not trying to describe all details of the brain or Artificial Neural Networks.
A fundamental feature of all living cells is the membrane potential which is an electric potential difference between the inside and outside of the cell. The potential difference is used as a source of energy to produce work. A special class of cells is excitable cells that can change their membrane potential to signal or change function. Neurons are excitable cells. They are considered the brain’s main building block. I hope you will partly disagree with the previous statement after this series of articles.
The brain consists of about 86 billion neurons, and each neuron has three sections — a cell body or soma, dendrites, and an axon.

Almost all neurons are multipolar in the central nervous system, which consists of the brain and spinal cord. Multipolar neurons have several dendrites emerging from the cell body. The dendrites receive synaptic signals from other neurons. Lastly, the cell body gives rise to one axon. The axon synapses with another cell, for example, a dendrite from another neuron, and conducts a signal.
Resting State
To understand how a signal is generated, we first need to understand the inactive or resting state. In a resting state, the cell has a potential difference of about -70mV. This results from the intracellular fluid having more negative anions than positive cations compared to the extracellular fluid, which has more cations than anions. As a consequence, an electric field is established across the cell membrane.
This ion imbalance is created through two mechanisms — the Sodium-Potassium-pump and ion-specific channels.
The Sodium-Potassium pumps create a chemical imbalance by extruding three Sodium ions and simultaneously importing two Potassium ions. As a result, the intracellular space has vastly more potassium ions than the extracellular space, while there are many more Sodium ions in the extracellular space compared to the intracellular space. Because this exchange happens against the chemical gradient of the ions, Sodium-Potassium-pumps need energy in the form of ATP to perform that action.
Ion channels allow for the passing of specific ions through the cell membrane. Neurons have an abundance of open Potassium ion channels in the resting state.
Assume, neurons would only have Potassium ion channels. In that case, Potassium ions would pass the channel from the intracellular space to the extracellular space following their chemical gradient created by the Sodium-Potassium-pump. However, since the channels only allow the cations to pass, the electric potential difference would increase, meaning the cell would get even more negatively charged. This creates an opposing electric force, attracting the positively charged Potassium cations back into the cell. Hence there is an equilibrium where the outward chemical gradient matches the inward electric attraction. This equilibrium can be calculated using the Nernst equation.

The neuron has more than one significant cation imbalance. In particular, there are vastly more Potassium cations in the cell, whereas there are many more Sodium cations and Chloride anions outside the cell. Using the Nernst equation, we can calculate the equilibrium potential for each ion.

The electric force is independent of the chemical compound creating the potential difference. Consequently, when combining several ions, we cannot just calculate the average of the Nernst equilibrium to calculate the membrane potential. We need to consider also the specific permeability. A particular ion with a high permeability will influence the aggregate membrane potential much more towards its own Nernst equilibrium than an ion with very low permeability.
This is taken into account in the Goldman equation, which essentially combines the Nernst equation for all three ions while accounting for their specific permeability.
Since ions are surrounded by polar water molecules, they cluster into large groups that cannot pass the bilipid membrane of the neuron. The specific permeability of each ion is consequently dependent on the number of specific ion channels. There are about 20 times more Potassium channels than Sodium channels. There are also about two times more Potassium channels than Chloride channels.

Using the specific permeabilities, we get a calculated potential difference of approximately -70mV, which matches laboratory measurements.
Temporal Summation
I will elaborate in a following article on how a cell is excited by receiving a signal from a presynaptic cell. For this article, let us take for granted that the neuron will change its state to excited after the cell membrane potential is changed from -70mV to -55mV. Passing this threshold is referred to as action potential. Through a mechanism elaborated in yet another article, the neuron passes a signal along the axon to the next cell or cells. If the threshold is not passed, referred to as a graded signal, no signal is conducted along the axon to the next cell or cells.
Since achieving the resting state through the mechanism outlined above takes about 5 ms, rapid graded signals cause a summation if not enough time (less than 5 ms) has passed to reestablish the resting potential. This phenomenon is called temporal summation and is unique to biological neurons.
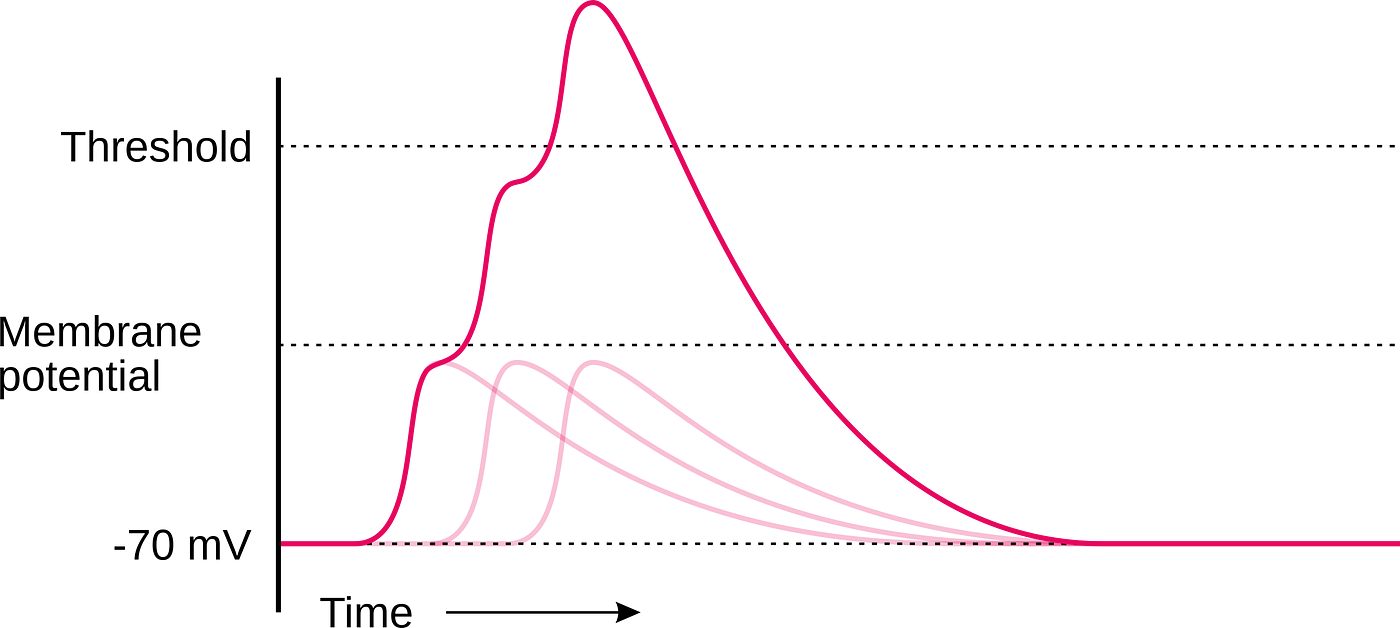
Temporal summation is a critical difference compared to artificial neurons. Several artificial neurons would be necessary to model temporal summation, compared to just one biological neuron.
Adding a working memory to artificial neurons is one of many approaches researched and tested. The working memory would model the time required to reestablish the resting potential using a decay function.